HD的主要发病机制是肠道末端的一段肠壁中神经节细胞缺失,导致该段肠管持续异常收缩,而临近的上段结肠仍然保持蠕动功能,结果导致上段结肠代偿性扩张、肥厚,最终形成巨结肠,因此该病又称无神经节细胞症[7]。
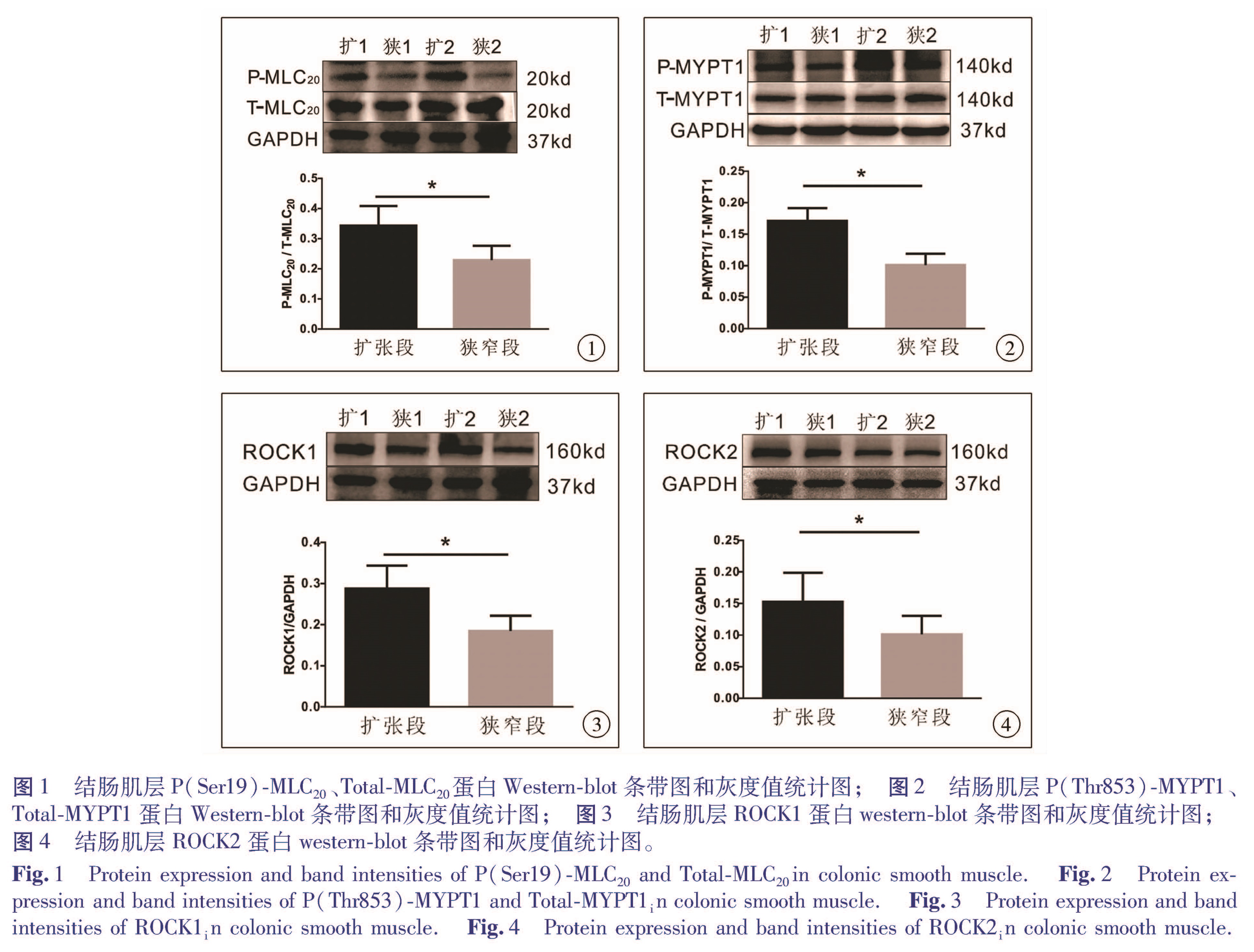
图1 结肠肌层P(Ser19)-MLC20、Total-MLC20蛋白Western-blot条带图和灰度值统计图; 图2 结肠肌层P(Thr853)-MYPT1、Total-MYPT1蛋白Western-blot条带图和灰度值统计图; 图3 结肠肌层ROCK1蛋白western-blot条带图和灰度值统计图; 图4 结肠肌层ROCK2蛋白western-blot条带图和灰度值统计图。
Fig.1 Protein expression and band intensities of P(Ser19)-MLC20 and Total-MLC20 in colonic smooth muscle.Fig.2 Protein expression and band intensities of P(Thr853)-MYPT1 and Total-MYPT1 in colonic smooth muscle.Fig.3 Protein expression and band intensities of ROCK1 in colonic smooth muscle.Fig.4 Protein expression and band intensities of ROCK2 in colonic smooth muscle.
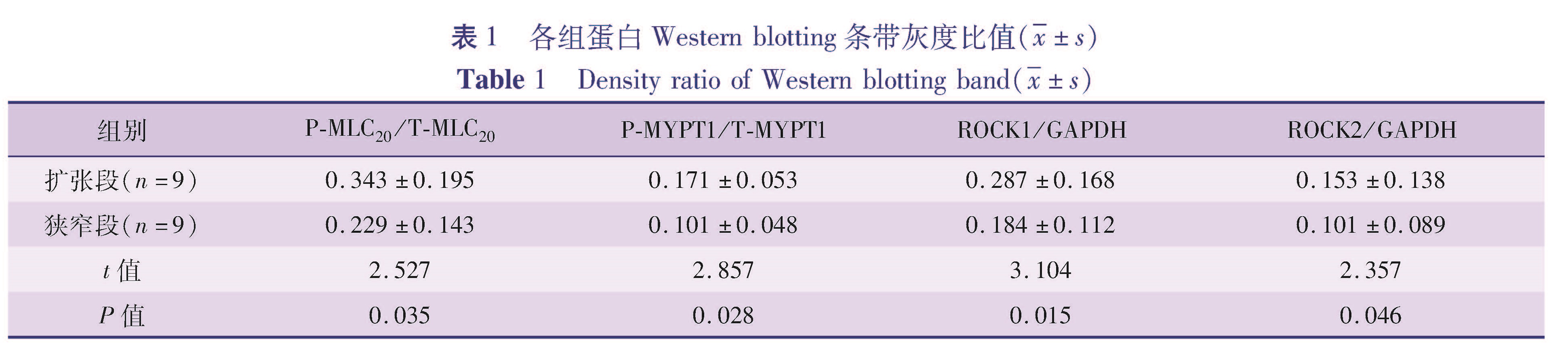
表1 各组蛋白Western blotting条带灰度比值(x^-±s)
Table 1 Density ratio of Western blotting band(x^-±s)
近20年来关于巨结肠的研究中,许多学者致力于探究ENS及消化道起搏细胞ICC。其中ENS释放多种神经递质,通过神经递质直接或者间接作用于平滑肌细胞,从而影响其收缩。在HD患者的病变肠段中,目前研究认为存在多种神经递质的释放异常,如肾上腺素能神经递质释放减少、一氧化氮合酶神经递质释放减少、胆碱能神经递质释放增多等[8]。这些神经递质的改变均通过影响平滑肌的收缩,最终对胃肠蠕动产生影响。另一方面,ICC不仅作为起搏细胞,将自发产生的节律性电兴奋传给临近的平滑肌细胞,使其去极化并产生慢波和快波,引起平滑肌收缩,使胃肠道产生节律性运动,也介导了神经信号的传递[9]。在多种消化道动力障碍性疾病的发生过程中,ICC细胞形态和功能的改变起到了关键性作用,其发育异常可能是一些儿童胃肠道疾病如先天性巨结肠发病的重要因素[10]。
此外,随着对PDGFRα+细胞形态和功能研究日益深入,2012年Sang Don Koh [11]首先在消化道平滑肌兴奋性的研究中提出了SIP合胞体(平滑肌/ICC/PDGFRα+细胞)的概念,认为胃肠道平滑肌的节律性收缩是外来神经、肠内神经丛与SIP合胞体协调作用的结果,即肠运动神经释放神经递质作用于肌层中ICC、PDGFRα+细胞这两种间质细胞,并且通过间质细胞与平滑肌细胞之间的缝隙连接进一步影响平滑肌运动[12]。也有越来越多的研究表明,在HD患儿无神经节细胞的肠段中,PDGFRα+细胞的功能发生了变化[12,13]。
综上所述,无论是ENS,还是SIP合胞体,最终都是通过影响平滑肌收缩来影响胃肠运动。衡量平滑肌收缩程度的一个重要因素即是肌球蛋白轻链(MLC20)的磷酸化水平。一般而言,结肠肌层的平滑肌细胞在受到外界刺激时,主要是通过“钙依赖”途径和“钙敏化”途径影响MLC20的磷酸化(图5)。“钙依赖”途径的大致过程是,在平滑肌细胞受到刺激后,胞外的Ca2+通过L-型钙通道进入胞内,与钙调蛋白结合,激活肌球蛋白轻链激酶(MLCK),促使MLC20磷酸化增加,平滑肌细胞发生收缩反应; 而“钙敏化”途径主要是RhoA/ROCK信号通路,外界刺激通过胞膜上的G蛋白偶联受体,经RhoA及其底物ROCK的介导,促使肌球蛋白轻链磷酸酶(MLCP)的亚基MYPT1发生磷酸化使MLCP自身的活性被抑制,MLCP的活性被抑制后可抑制MLC20的去磷酸化过程,间接增加MLC20的磷酸化水平,促进平滑肌收缩[11,14]。这两条途径相互协调,最终完成平滑肌的收缩反应。然而,目前在HD发病机制的研究中,ENS、ICC细胞和PDGFRα+功能的改变,对肌层平滑肌细胞自身产生了怎样的影响,尤其是与这两条途径相关的影响,少有研究报道。近年来有研究发现,在HD无神经节细胞肠段中,肌层的平滑肌细胞存在层粘蛋白、结蛋白、纽蛋白、营养不良蛋白和εSG的缺失,smoothelin蛋白表达减少,这些蛋白主要在平滑肌细胞的增殖分化阶段发挥作用[15]。同时Won KJ等[16]的研究指出,HD大鼠巨结肠病变部位相较于正常大鼠结肠对应的部位,其肌层厚度增加明显,可能存在平滑肌细胞过度增殖。此外,周小琴等[17]对HD患儿病变段肌层组织进行α-SMA免疫染色发现,病变段平滑肌细胞α-平滑肌肌动蛋白(α-SMA)染色相对于正常肠段染色较浅,α-SMA表达减少。α-SMA是由成熟平滑肌细胞表达的与肠蠕动密切相关的收缩蛋白之一。这些均提示HD病变肠管的平滑肌细胞本身可能存在发育和功能的异常。而在本实验中,我们发现9例HD患儿狭窄段的肠壁肌层中,与RhoA/ROCK信号通路相关的蛋白表达(ROCK1,ROCK2,P(Thr853)-MYPT1)均比自身扩张段减少。说明在HD患儿狭窄段的平滑肌细胞中,相较于扩张段,RhoA/ROCK信号通路反应性是减弱的,这与在大鼠模型中观察到的结果相反[6]。ROCK包括ROCK1和ROCK2两种亚型; ROCK1主要分布于肝、肾、肺等组织器官中,与组织细胞的生长和凋亡相关; ROCK2主要在肌肉组织和脑组织中表达,其在MLCP介导的细胞骨架构建过程中发挥重要的作用[18]。可见,在平滑肌细胞正常的收缩过程中,主要是ROCK2参与“钙敏化”途径。而我们的结果表明,在HD狭窄段,ROCK1和ROCK2的表达均是降低的。Coyle D等[19]研究发现,在HD患儿无神经节细胞的肠段中,ROCK1的表达减少,ROCK2蛋白未发生明显改变,这和我们的研究结果部分一致,说明在HD患儿病变肠段中,ROCK的功能可能也发生了变化。
另一方面,有研究表明在HD患儿无神经节细胞的肠壁中,肌层平滑肌细胞表面的L-型钙通道的表达、电流强度和电流密度与有神经节细胞的肠段相比,并没有明显区别[20]。L-型钙通道是胞外Ca2+进入胞内,引起平滑肌细胞通过“钙依赖”途径收缩的重要通道,说明“钙依赖”途径在无神经节细胞的肠壁中可能并没有受到影响。且有研究指出,在病变肠段中,存在多种神经递质释放异常,主要是兴奋性神经递质释放增多[7]。乙酰胆碱(Ach)是胃肠道最主要的兴奋性神经递质,由于胆碱能神经纤维的增生,Ach释放增多[21]。释放的Ach可促使胞外的Ca2+通过L-型钙通道进入平滑肌细胞内,引发“钙依赖”途径,促使平滑肌细胞收缩。因此,这些均可能是HD病变段表现出痉挛状态的原因。
本研究表明,HD患儿肠段中,相对于扩张段,狭窄段肌层平滑肌细胞RhoA/ROCK信号通路减弱,“钙依赖”途径较强,提示在结肠平滑肌中“钙依赖”途径和“钙敏化”途径存在功能失衡,这可能是HD无神经节肠段平滑肌收缩异常的内在机制。