排便是由中枢神经、周围神经以及结直肠平滑肌和肛门括约肌等共同参与的一个复杂过程,其中枢神经包括脊髓内的低级中枢以及颅脑内的高级中枢[1-2]。此外,排便与排尿过程也有着紧密的联系,与二者相关的组织器官不仅在解剖位置上紧密相邻,而且在神经反射通路方面也较为相似。这就要求在示踪排便反射神经通路时,所选择的神经示踪剂以及示踪方法既要保证示踪过程的特异性,避免示踪剂局部扩散出现假阳性的结果,又要保证示踪剂能够跨越多级神经元最终到达颅脑内的高级中枢,从而完整反映出整个神经通路的概况。而一般的神经示踪剂如辣根过氧化物酶,荧光染料等均无法满足以上要求[10],故本次实验选用了伪狂犬病毒的减毒株(Bartha株)作为神经示踪剂。它作为一种嗜神经病毒,能够在神经元细胞内自我复制,从而保证了示踪过程中有足够浓度的报告蛋白以显示整个神经通路而不会出现信号衰减的现象; 此外,它还能够跨越突触传播至与其有突触连接的神经细胞,而不会扩散到周围无突触连接的神经元,从而保证了其示踪过程的特异性; 最后该病毒的减毒株,在大大削弱了病毒毒力的同时又保留了其逆向跨突触传播的能力,从而延长了被感染动物的存活时间,使病毒有足够的时间来逆向感染示踪整个与靶器官相关的神经通路。该病毒不仅能够较为完全的显示与排便反射相关的整个神经通路,而且能够保证示踪结果的特异性,是一种较为理想的神经示踪剂[3-4]。但其最后示踪结果的检测仍然需要免疫组化染色,而影响免疫组化染色结果的各种因素势必也会对示踪结果产生较大的影响,这也是本次实验的不足之处。近期一些伪狂犬病毒的基因重组体如PRV — 152和PRV — 614,分别标记了GFP(绿色荧光蛋白)基因和RFP(红色荧光蛋白)基因的出现,使得病毒的示踪结果可以在荧光显微镜下直接观察而无需进一步染色处理,从而增加了神经传导检测的可靠性,以后将会得到广泛的应用[11]。
目前,普遍认为控制排便的低级中枢位于脊髓的腰骶段,且包括本次实验在内的多项研究均证实该反射通路与脊髓L6至S1节段关系密切。更有学者认为与L6节段相比,排便反射的传入与传出神经更多的起源于S1节段,这也为骶副交感神经核团控制盆腔脏器功能的学说提供了依据。在本次大鼠实验中不仅在腰骶段,同时在胸腰段脊髓的T13至L1节段发现了大量伪狂犬病毒阳性的神经元,这可能与胸腰段的交感神经节及内脏神经向盆腔内所投射的交感神经纤维有关[7]。而控制排尿、排便以及生殖功能的交感/副交感神经通路在功能与解剖上紧密相连,仍需要进一步的研究以明确区分。
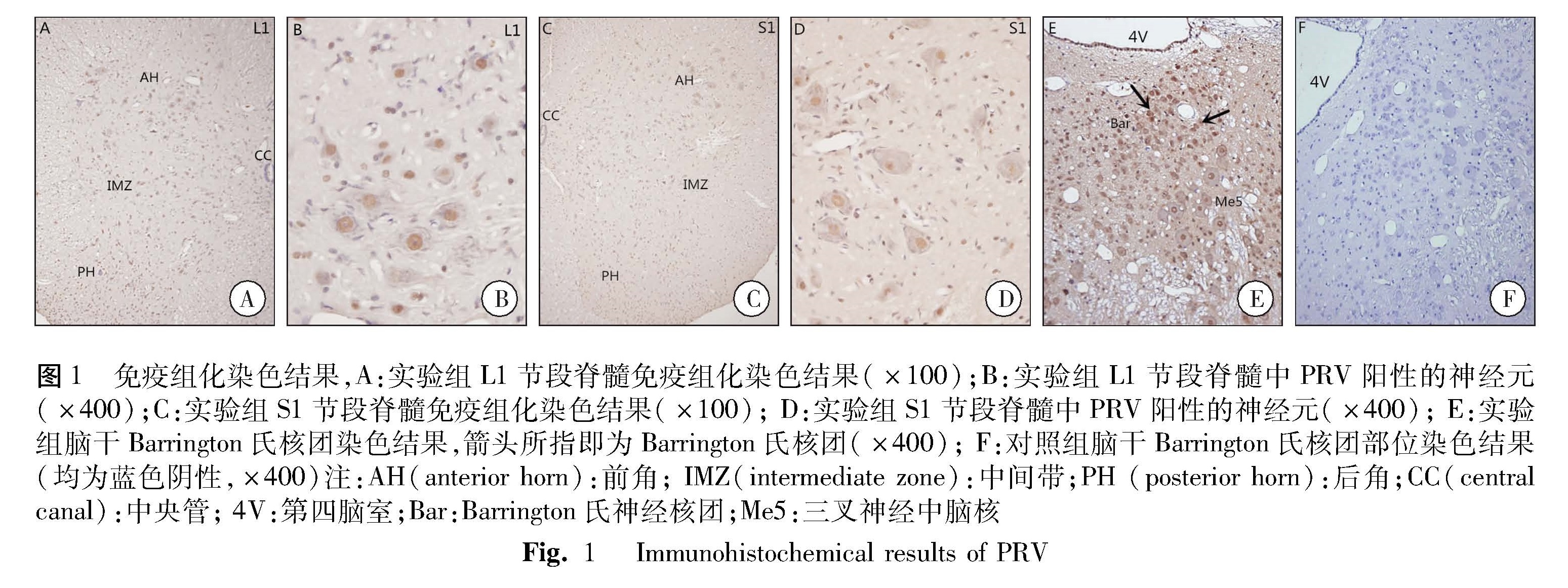
在本次试验中我们在大鼠脑干的Barrington氏神经核团内发现了大量伪狂犬病毒阳性的神经元,说明这之前被认为是脑干排尿中枢的神经核团在排便的控制方面也起着重要的作用[5,7-8]。也有学者使用在膀胱和结肠进行双标记的方法进一步证实在该核团内至少存在三块区域,即分别控制排尿与排便的相对独立的区域,以及共同调控二者功能的区域[8]。而这一功能区域划分的详细情况仍需要进一步研究。此外,由于动物和人类排便控制条件的不同,本次实验并未涉及伪狂犬病毒向大脑皮层内的示踪结果,而排便反射神经通路在高级中枢神经系统内的部分,对研究某些与排便相关的先天性畸形如先天性巨结肠、先天性肛直肠畸形等疾病的发病原因、诊疗方法以及预后情况等方面具有至关重要的作用,这也是今后需要努力研究的方向[12-13]。